LA HIBRIDACIÓN Y LA TRANSMUTACIÓN
Un tema muy comentado y que suscita muchas preguntas y pólemica es el tema de la hibridación, para ello expondre varias de las preguntas frecuentes y su respuestas, espero que os sean de ayuda.Primero haremos un esquema de las especies con anillo y sin anillo ocular ( también llamado perioftálmico).
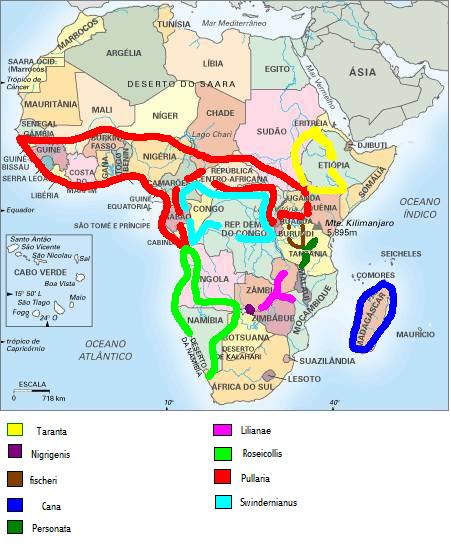
Con anillo ocular tenemos;
-Agapornis personata.Fue presentado a la ciencia por Reichenow en 1887.
-Agapornis fischer.Fue presentado por Reichenow en 1887.
-Agapornis lilianae.Fue presentado a la ciencia por Shelley en 1894
-Agapornis nigrigenis.Presentado por Sclater, WL, en 1906.
Sin anillo ocular;
-Agapornis cana. Tenemos dos subespecies; agapornis cana cana, presentado a la ciencia por Gmelin en 1788, y agapornis cana ablectanea por Bangs en 1918
-Agapornis roseicollis.Tenemos dos subespecies, roseicollis roseicollis, presentado a la ciencia por Vielliont 1917 y roseicollis catumbella por Hall en 1952
-Agapornis taranta.Tenemos dos subespecies, taranta taranta presentado a la ciencia por Stanley en 1814, y taranta nana por Neumann en 1931
-Agapornis swinderniana.Tenemos 3 subespecies, agapornis swindernianus swindernianus( Kuhl, 1820), agapornis swindernianus emini ( Neumann, 1908) y agapornis swindernianus zenkeri ( Reichenow,1895)
- Agapornis pullaria.Tenemos 3 subespecies, agapornis pullarius pullarius (Linnaeus, 1758), agapornis pullarius guineensis ( Statius Müller, 1776), agapornis pullarius ugandae (Neumann, 1908).
Mucha gente hibrida por desconocimiento, porque les venden en las tiendas pájaros que ni ellos mismo saben qué son, por informaciones erróneas y por estar mal aconsejados. Por lo que debemos estar informados si queremos tomarnos en serio esta afición y queremos disfrutar de la belleza de los agapornis, ya sabeis nadie nace sabiendo, por lo que lo mejor es preguntar mil veces lo que no se sepa y leer mucho y aprender de los criadores mas experimentados y que de verdad sepan de lo que estan hablando.
¿Qué especies al cruzarlas dan descendencia fértil?
Si cruzamos especies de anillo ocular( lilianae, nigrigenis, personata y fischer) estas crias serán fértiles y no seran ni una cosa ni otra, estos pájaros al cruzarlos nuevamente daran híbridos y todos los cruces posteriores segurian dando ejemplares fértiles.
Hay también gente que habla de cruces de taranta con roseicollis los cuales carecen de anillo y tambien serían fértiles, pero yo como no tengo constancia segura de ello no lo desarrollare más.
¿Qué especies al cruzarlas dan descendencia infértil?
Cuando cruzamos especies de anillo acular con no ocular(roseicollis, tarantas..)la descendencia será infértil y estos pájaros no podran tener descendencia, esto no quiere decir que no tengan instinto reproductor, ya que incluso se aparean y ponen huevos como los demás, pero los huevos son infértiles, ya los crucemos con otros infértiles o con ejemplares puros no habra fecundación, y por no tanto no habra embrión.
Mención aparte merecen los comentarios de que estos cruces no generan agapornis estériles en su totalidad, y que estos hibridos necesitan más tiempo e incluso varios años para madurar y criar, yo personalamente no tengo constancia visual de ello, se comenta que es cierto, pero en todo caso no hagais la prueba.
Después de estas explicaciones quiero decir desde aquí que no debemos cruzar especies, ya que algunas de ellas son dificiles de encontrar puras y algunas de ellas son escasas en su hábitat natural.
¿Qué es una transmutación?
Cuando se quiere pasar un color determinado de una mutación de una especie a otra, hablamos de transmutación, y se hace entre ejemplares que dan descendencia fértil evidentemente, como ejemplo, la mutación lutino surgio por primera vez en los agapornis lilianae en Australia sobre el 1936, y como esta mutación no existía en las demás especies se cruzaron ejemplares lutinos lilianae con personatas para pasar esa mutación a los personata, y de los personata se paso posteriormente a los fischer. Lo mismo paso con la mutación azul que se paso a través de transmutación de personata a fischer.
Estas transmutaciones deben hacerlas criadores expertos y no cualquiera que quiera experimentar.Por lo tanto transmutar es hibridar entre ejemplares que dan descendencia fértil de forma controlada y por expertos en la materia.
Para trasmutar se debe tener una excelente línea pura para que cuando hagamos la transmutación sucesivamente se vayan emparejando estos ejemplares que son hibridos e ir acercándolos hacia el standard o color puro, por lo que este proceso conlleva muchos años de selección y lo peor de todo son esos ejemplares intermedio(hibridos) que quedan disponibles.
Os preguntareis qué ocurre con esos pajaros, hay dos versiones, sacrificar a todos esos pajaros intermedios para asi no dañar las líneas de las especies puras o lo que yo creo que se suele hacer, que es venderlos como puros llenando el mercado de hibridos.
Un criador honesto que transmuta debera informar a sus compradores de lo que realmente cria,pero no siempre es así, tambien se da el caso de criadores que realmente no saben que sus pajaros no son puros ya que algunas mutaciones son dificiles de distinguir, y los venden sin mala intención,pero perjudicando a la especie y haciendo mas difícil la tarea de conseguir ejemplares puros.
Bajo mi punto de vista sacrificar a las crias para conseguir un tipo de mutación a base de manipular y matar aves no lo veo correcto,pero se hace y todos disfrutamos de estos colores sin pensar en ello, ahi cada cual tiene su criterio y no profundizare en él.Aunque como ya he dicho anteriormente yo creo que la mayoria de estos hibrídos los venden a importadores y pajarerias.
Cuando una mutación se pasa de una especie a otra hasta la sexta generación no podemos decir que tenemos pájaros puros de esta mutación, cuando el porcentaje de la pureza es superior al 98%, Ahora hare un pequeño esquema de una trasmutación:
- fischer puro x personata puro = personata-fischer (50% sangre fischer y 50% sangre personata).F1
- fischer puro x F1 = personata-fischer ( 75% sangre fischer y 25% sangre personata )F2
- fischer puro x F2 = personata-fischer ( 87,5% sangre fischer and 12,5% sangre personata)F3
- fischer puro x F3 = personata-fischer ( 93,75% sangre fischer y 6,25 % sangre personata)F4
- fischer puro x F4 = personata-fischer ( 96,87% sangre fischer y 3,13% sangre personata)F5
- fischer puro x F5 = personata-fischer ( 98,43% de sangre fischer y 1,57% sangre personata) F6
Y para alcanzar estos porcentajes de pureza de sangre tenemos que estar seguros que el fischer que cruzamos con el primer personata y con el persofischeri (F1, el F2…)son puros y ya sabemos lo dificil que es esto.
Los F son ejemplares intermedios( hibridos) y conforme los cruzamos con puros vamos adquiriendo pajaros mutados con más porcentaje de sangre fischer , asi se hacen cruces sucesivamente hasta la quinta o sexta generación que es en la cual se supone que obtenemos ejemplares con sangre fischer por lo menos superiores al 98%, por lo que los pajaros transmutados mantienen algunos signos minimos de sangre personata( por ese 1,57%) ,que se podran mostrar en su descendencia y los cuales debemos trabajar en nuestros pájaros.
Para mi un pájaro mutado puro es un ave que siendo mutada mantiene caracteristicas similares con el ancestral, como ejemplo los fischer azules, deben de tener la eumelanina de la cabeza igual que un ancestral bueno ya que lo que se elimina es la psitacina y la eumelanina debe queda intacta, y no tener la cara mancha con intromisión del negro, ni una linea horizontal por detrás de la nuca signo de la trasmutación con personata, deben de tener una linea diagonal por detrás de la cabeza que es lo que se exige en concursos.
Si queremos hacer la prueba podemos cruzar nuestros pájaros mutados que consideremos buenos con un ancestral( verde)bueno, si la descendencia se aleja mucho del ancestral entonces nuestros pájaros no son todo lo puros que pensabamos y se debería trabajar mas esa línea, cruzándolos con verdes puros, tarea dificil y que lleva años de dedicación, si por el contrario la descendencia es como uno puro ancestral, tendremos pajaros buenos y con esos se trabajara mejor.
Por último desde aqui quiero decirle a la gente que este leyendo mi artículo que no hibride, que deje las transmutaciones para gente experta, ya que el resultado casero de hibridar no nos lleva a ningún lado ni supone un gran mérito para el criador aficionado ya que es fácil conseguirlo(hibridar entre dos especies), y mediante esto no vamos a descubrir mutaciones nuevas, solo pájaros intermedios que en el caso de ser fértiles, nos haran mas difícil la tarea de conseguir pájaros puros y que ensucian nuestra afición.
Aurora Carranza Román