EVOLUCIÓN DE LOS AGAPORNIS
La primera tentativa para entender la evolución del género Agapornis fue hecha por Moreau (1948), que resumió lo que era sabido sobre la distribución, la ecología, la morfología, y el comportamiento de los agapornis. Usando toda esta información, él propuso una clasificación del género. Moreau( 1948)agrupo los agapornis en base a siete morfológicos y caracteres del comportamiento.
Asi en líneas generales tenemos estos dos grupos;
GRUPO A: cana, pullaria, taranta. Los llamados primitivos.
GRUPO INTERMEDIO; roseicollis.
GRUPO B: fischeri, personata, lilianae, nigrigenis.Los llamados avanzados.
Grupo A ;
- Plumón al nacer blanco.
- Plumaje distinto del juvenil ;presente.
- Piel blanca alrededor del ojo; ausente.
- Dimorfismo sexual;presente
- Barra negra en plumas centrales de la cola ;presente
- Método de llevar el material de nidificación; en las plumas
- Forma del nido; relleno
Grupo intermedio;
- Plumón al nacer; rojo.
- Plumaje distinto del juvenil; presente.
- Piel blanca alrededor del ojo;ausente.
- Dimorfismo sexual; ausente.
- Barra negra en plumas centrales de la cola; ausente.
- Método de llevar el material de nidificación; en las plumas.
- Forma del nido; copa
Grupo B;
- Plumón al nacer; rojo.
- Plumaje distinto del juvenil;ausente.
- Piel blanca alrededor del ojo; presente.
- Dimorfismo sexual;ausente.
- Barra negra en plumas centrales de la cola;ausente.
- Método de llevar el material de nidificación;en el pico.
- Forma del nido; cúpula.
La mayor parte de nuestro conocimiento del comportamiento de los agapornis es resultado de observaciones de agapornis en cautividad hechos por Dilger (1960, 1961). Sus estudios incluyeron a todas las especies de Agapornis a excepción de swinderniana , y ellos proporcionaron descripciones de la crianza y del comportamiento social. Y por otro lado los estudios que se han hecho posteriormente utilizando pruebas de ADN.
De acuerdo con sus observaciones del comportamiento, Dilger (1960, 1961) caracterizó a los cana, taranta, y pullaria como “primitivos,” y fischeri, personata, lilianae, nigrigenis “los más desarrollados El comportamiento de la especie restante, roseicollis, parecía intermedio, conduciendo Dilger a concluir que se presentó de la especie “primitiva” y es ancestral a la especie “altamente desarrollada” (Dilger 1960, 1961).
Dilger conicidió con Neunzig (1926) y Hampe (1957)que fischeri, personata, lilianae, y nigrigenis son probablemente subespecie de una especie. Él también sugirió que el taranta y el pullaria estén relacionados muy de cerca el uno al otro que con las otra especies del género, y que son los parientes más cercanos del cana, que es endémico de Madagascar.Aunque él generalmente estaba de acuerdo con el postulado de Moreau, Dilger (1960: 650) sugirió que las especies del grupo B estan relacionadas de cerca con los roseicollis.
Cana y taranta nidifican en agujeros de árbol, y Pullaria utiliza las cavidades excavadas en los termiteros .En las tres especies, la cavidad del nido se teje con material ( cáscaras de semilla, pedazos pequeños de la corteza, hierba u hojas) que la hembra lleva remetido en sus plumas del cuerpo . Los hábitos de nidificación de una cuarta especie, swinderniana ,se conocen poco pero se cree que puede nidificar en termiteros también .
Las hembras de los roseicollis llevan el material de nidificación (tiras de corteza, de hojas, o de hierba) remetido sus plumas de la rabadilla y construyen nidos en forma de plato o taza dentro de las cavidades.
Las hembras de las cuatro especies restantes (fischeri , personata , lilianae .y los nigrigenis ) llevan el material de nidificacion (los tallos y tiras largas de corteza) en sus picos y construyen nidos abultados y abovedados dentro de las cavidades. El material de nidificacion se teje todo junto, y la estructura que resulta conserva su forma aunque se saque de la cavidad .
La designación de algunos como “primitivos” y de otros como “avanzados” resulto de la selección de Loriculus como el género lo más relacionado cercanamente posible , y porque los miembros del primer género también llevan el material de nidificaion en sus plumas y son sexualmente dimorficos.
Los roseicollis parecen ser intermedios con respecto al “primitivo” (grupo A) y avanzado (grupo B) porque comparte tres caracteres con cada grupo. Colocandose como un intermedio filogenico indicaría que el comportamiento de construccion de los nidos se desarrolló del hábito de tejer los nidos, reflejando una elaboración gradual de la construccion de los nidos.
Sin embargo, Moreau (1948: 236) sugiere que los roseicollis y las especies del grupo B(avanzados) son distintos linajes: ....... es curioso que de todos los pájaros del grupo B, el agapornis nigrigenis siendo el miembro que geográficamente es más cercano a los roseicollis , es el menos parecido a el. Esto sugiere que, aunque los roseicollis son intermedios en caracteres,el Grupo B es mas desarrollado.
Todas estas especies primitivas(grupo A) utilizan las cavidades en la naturaleza para construir sus nidos, tejiendo con material la cavidad muy poco, sin embargo, ocasionalmente algunos construirán un nido más estructurado. Estos pájaros construyen mejor en cajas verticales, horizontales y diagonales, aunque las diagonales y las horizontales parecen ser las preferidas. Son para criadores experimentados. El pullaria es el más difícil, a menudo requiriendo una espuma o corcho alrededor del interior de la caja. El cavar en el nido al parecer impulsa su instinto de crianza.
Los roseicollis son el agaporni (intermedio) más común, construirán un nido en forma de tazon ya sea de papel, de paja, de hierbas o de otro material. Es relativamente fácil conseguir que crien y es un placer verlos, aunque es ruidoso y agresivo. No parece importarle que lo molesten tanto como las tres especies anteriores.
Las especies avanzadas incluyen al personata, Nigrigenis, Lilianae y fischeri. Todos construyen los nidos en forma de cúpula usando flexibles ramitas de arboles. Nidifican en la cavidad con estos materiales, y a menudo lo hacen tan complicado que resulta difícil al criador mirar adentro y determinar el desarrollo de la nidada.
Los nigrigenis son las especies más avanzadas e incluirán un compartimento falso sobre el compartimento verdadero del nido. Proveer a estos agapornis mas tranquilos (los nigrigenis d y los lilianae ) del material correcto y gozarás de los sonidos más suaves que emiten
La relación levemente más cercana entre los nigrigenis y los lilianae primero fue observada por Moreau (1948), y esta relación es apoyada por los datos del mtDNA ( adn mitocondrial).
Segun Moreau , la separacion entre los clanes nigrigenis/lilianae y personata / fischeri probablemente fue causada por las montañas en la cabeza del lago Nyasa.
El final norteño de la división de personata/fischeri coincide con la localización del Rift Valley(valle de la grieta), que puede ser responsable de su separación; sin embargo, en el final meridional de la división, ningunas barreras geográficas importantes separan a miembros dentro de los dos pares de la especie (Moreau 1948).Moreau (1948) sugirió que la separación es debida por las regiones del arbolado del bosque “miyombo”, y, en el caso de nigrigenis y de lilianae, por una región de la alta elevación.
La filogenia del mtDNA también demuestran una relación cercana entre el taranta y el pullaria, que fue observado por Moreau (1948) y Dilger (1960)con anterioridad.
La posición básica del cana indica que probablemente se aisló en Madagasca relativamente temprano en el curso de la diversificación del género Agapornis. Sin embargo, la cantidad de divergencia de la secuencia entre el cana y sus parientes más cercanos del continente (taranta y pullaria) sugiere que la colonización de Madgascar ocurrio después de la separación de la isla del continente África.
Observamos que dentro delos Agapornis, hay una correlación entre la adopción de los nidos y la construcción de losmismos: los roseicollis y los lilianae utilizan con frecuencia los nidos del tejedor (Forshaw 1989), y el fischeri y el personata parecen utilizar los nidos de otros pájaros (Moreau 1948).
Esto esta relacionado con la hipótesis de la nidificación indicando una progresión del comportamiento hacia el tejer nidos más elaborados.
La construcción de un nido dentro de una cavidad puede ser ventajosa porque permite a los pájaros modificar las cavidades que de otra manera serían inadecuadas para criar (Eberhard 1997).
Vriends (1978) encontró que si la abertura del nidal es demasiado grande, los pájaros a veces apilan el material de nidificacion uno encima del otro para hacer la entrada más pequeña. Él también observó que el grado de la construcción del nido depende del tamaño de la cavidad, si el espacio es limitado, la bóveda puede ser omitida. Vriends también observó que las hembras agregarán material adicional a sus nidos si hay escapes de material en el nido, o cuando hay algunos pinchones en el interior del nido.
Una diferencia importante entre las especies de Agapornis que construyen nidos y las que no lo hacen es criar en colonia. Los que construyen nidos (anillados y los roseicollis) son coloniales mientras que los otros son criadores solitarios.
En cautiverio, las parejas de cana y de taranta criando son extremadamente territoriales y se deben mantener las jaulas separadas (Vriends 1978, Erhart 1983).
La crianza gregaria (colonial) se pudo haber facilitado por la evolución de la forma de construir los nidos, porque la capacidad de construir un nido podría dar a las parejas de cria flexibilidad en la eleccion de los sitios de nidificacion (Eberhard 1997).
Una asociación entre la construcción cada vez más compleja de nidos y el anidamiento gregario también se ha demostrado en un análisis filogenetico de la evolución de la construcción de la jerarquía en golondrinas(Winkler y Sheldon 1993). En el caso de las golondrinas, Winkler y Sheldon sugieren que la transición de nidos de la taza a nidos abovedadas permitio criar mas pichones.
En Agapornis, el cambio de dimorfismo sexual (en la especie “primitiva”) al monomorfismo sexual (en roseicollis y anillados) puede ser un resultado del cambio a la crianza colonial (Dilger 1960), quizás debido a la selección no sexual por el color, sino social.
Uno de éstos es el método de llevar el material de nidificación; el cambio de llevar material en las plumas a llevarlo en el pico permite transportar material más grandes y/o más pesados.
Los cambios del plumaje se pudieron haber seguido de los cambios en comportamiento de edificar nidos, porque el interruptor del dimorfismo al monomorfismo se liga probablemente al cambio de la forma de criar en solitario a la crianza gregaria. Es probable que la crianza gregaria fue facilitada por la capacidad de construir las estructuras de nido dentro de las cavidades, porque la capacidad de modificar las cavidades inadecuadas daría a la pareja de cria la creciente flexibilidad en el sitio de anidar.
sábado, enero 06, 2007
viernes, diciembre 01, 2006
El GRIT EN PSITÁCIDAS; ¿es adecuado o no?
EL GRIT EN PSITÁCIDAS
¿Necesitan grit los loros? Ésta es una pregunta común que nos concierne a criadores y a dueños de agapornis como mascotas, pero también es una fuente de desacuerdo. Entender qué es el grit, su propósito, y los posibles problemas que pueden resultar de su uso, pueden ayudarnos en decidir si ofrecerlos o no a nuestros pájaros. La mayor parte de la información presentada aquí se ha recopilado y se ha resumido del libro Avian Medicine: Principles and Application by Ritchie, Harrison and Harrison.
Para empezar debemos hacer un pequeño recordatorio del aparato digestivo para así entender algunos conceptos posteriores.
APARATO DIGESTIVO
Se encuentra constituido por orofaringe, esófago, estómago, duodeno, yeyuno, íleon, un par de ciegos y colon. Este último desemboca en la cloaca, que es un segmento final también para el aparato urinario y genital. El hígado y páncreas secretan sus productos al intestino delgado. El sistema digestivo está diseñado para que se pueda ingerir la máxima cantidad de comida en el menor tiempo posible, para huir de sus depredadores.
Las psitácidas tienen más papilas gustativas que los otros pájaros, y están situadas en el paladar, faringe y base de la lengua.
El sentido del gusto de los loros puede ser comparado con el de los mamíferos, ya que pueden detectar partículas en el agua para no beberla.
1. Orofaringe ;Este término se aplica a la cavidad que va desde el pico al esófago, ya que las aves no poseen paladar blando y por tanto no existe división entre cavidad oral y faringe como los mamíferos. El techo de esta cavidad lo conforma el paladar, y el suelo la mandíbula, lengua y la elevación o prominencia laríngea.
No existen labios ni dientes, elementos que son reemplazados por el pico y el estómago muscular o molleja
2. Esófago ; Situado entre la tráquea y músculos cervicales, pero pronto se desvía hacia la derecha. En la entrada del tórax, la pared ventral del esófago se expande y forma el buche (ingluvium) ubicado hacia la derecha.
-El Buche almacena el alimento durante un periodo corto cuando el estómago muscular está lleno. El buche produce la leche ingluvial, que mezclada con alimentos ingeridos es regurgitada y administrada a los pichones, esta leche se produce por una descamación de células epiteliales cargadas de grasa de ambos sexos provocada por una estimulación de oxitocina y prolactina. Es rica en grasa y proteína pero carente en carbohidratos.,el buche no tiene glándulas digestivas. En psitácidas está orientado transversalmente en el cuello, en forma de S y cuando está distendido se cuelga de los músculos del pecho.
Luego el esófago pasa sobre la bifurcación de la tráquea, ventral a los pulmones y dorsal a la base del corazón, y continúa con el proventrículo o estómago glandular hacia la izquierda.
3. Estómago se encuentra constituido por dos secciones;
-la primera, llamada proventrículo, o estómago glandular cuya mucosa está tapizada por múltiples papilas, por donde desembocan los conductos de las glándulas, cuya secreción es rica en ácido clorhídrico, pepsinógeno y mucina, que es muy importante para el inicio de la etapa de digestión.
- la segunda, se conoce como molleja o estómago muscular, de mayor tamaño, sus paredes están formadas por fuertes músculos y su función es la de triturar los alimentos. Interiormente está tapizado por un grueso epitelio de color amarillo queratinizado que la protege de los posibles daños que pueden causar los guijarros o piedrecillas ingeridas.. Tiene forma discoide y su mucosa está protegida por un duro complejo polisacárido-proteico llamado KOILIN. El tipo y tamaño del grit en la molleja está en acordancia con la dieta y su función es triturar el alimento, supliendo así la función de los dientes. El grit no es esencial para una digestión normal y puede causar impactaciones.. Es común en las gallinas de campo que en el interior de este órgano se encuentren piedritas, que van a formar parte de una función mecánica, contribuyendo al triturado del alimento, supliendo así la función de los dientes.
4. Duodeno Se dispone en forma de asa duodenal dentro de la cual se dispone el páncreas y sus dos o tres conductos excretores que desembocan en el extremo distal del duodeno. Del hígado proviene dos conductos, el hepatoentérico y el hepatocístico que desembocan cerca de los pancreáticos.
5. Yeyuno Son asas móviles sostenidas por mesenterio. Presenta un pequeño apéndice o divertículo vitelino que era la antigua conexión del intestino con la vesícula vitelina (la vesícula vitelina persiste dentro de la cavidad corporal, después de la eclosión, para nutrir el pollito durante los primeros días). En la mucosa yeyunal se pueden observar placas generadas por nodulillos linfáticos agregados.
6. Ileon Continúa del yeyuno, sin clara demarcación considerando su origen a partir de los vértices del ciego.
7. Intestino Grueso Compuesto por los ciegos y el colon. Los ciegos son largos, originados en la unión ileocólica y distribuidos en forma retrógrada a lo largo del íleon; al que se unen por los pliegues ileocecales. El segmento proximal de los ciegos tiene una potente capa muscular o esfínter cecal con abundante tejido linfoide denominado tonsila cecal. En los ciegos se genera degradación bacteriana de la celulosa. Las aves psitácidas carecen de ciegos.
8. Colon De unos 10 cm de longitud termina en un ligero ensanchamiento de la cloaca. El coprodeo es la continuación ampuliforme(forma de ampolla) del colon donde se acumulan las heces, limitado caudalmente por el pliegue coprourodeal.
9. Cloaca Es un órgano común a los aparatos digestivo, urinario y genital y desemboca al exterior por el orificio cloacal externo. Desembocan a la cloaca, el colon, ureteres, conductos deferentes o el oviducto izquierdo en diferentes niveles. La cloaca se divide en tres regiones: - la anterior o coprodeo, que recibe el excremento del intestino- la intermedia o urodeo, que recibe las descargas de los riñones a través de los uréteres y a la vez es donde desembocan el oviducto o los conductos deferentes.- la posterior o proctodeo, que es la más grande y muscular. Su contracción expulsa violentamente los excrementos del ave.
Indicar también que la gran capacidad digestiva que presentan las aves hace que utilicen la mayor parte del alimento ingerido, por lo que sus excrementos, aunque numerosos, son de escaso tamaño.
10. Hígado y Páncreas El hígado está formado por lóbulos derecho e izquierdo, unidos cranealmente. De mayor tamaño el lóbulo derecho y en su cara visceral se encuentra la vesícula biliar que no está presente en palomas y algunas psitácidas. Este lóbulo derecho está perforado por la vena cava caudal. El lóbulo izquierdo está dividido.
11. Páncreas Es alargado situado en el asa duodenal formado por dos lóbulos, uno dorsal y otro ventral conectados distalmente. Posee dos o tres conductos que llevan el jugo pancreático al duodeno.
12. Bazo De forma esférica, de unos 2 cm, de color marrón rojizo.
¿Para qué se usa el grit?
El grit es utilizado por los pájaros para ayudar en la digestión de semillas enteras, intactas. Las enzimas digestivas de los pájaros digieren la porción interna de semillas, pero tienen dificultad de accion por la capa fibrosa(cáscara). Las cascaras de las semillas enteras, tales como semillas de girasol, actúan como una barrera, evitando que las enzimas digestivas alcancen el contenido nutritivo del interior de la semilla.
El grit en el ventrículo, ayuda en moler cáscaras y usar lejos las externas de las semillas, permitiendo a las enzimas digestivas alcanzar la porción interna valiosa. Si hay cualquier otro propósito para el uso de la arena por los pájaros, todavía no se sabe.
¿Qué es el grit?
El término grit se aplica a dos grupos de sustancias; arena insoluble y arena soluble.
- La arena insoluble, es el tipo que es discutido aquí, se compone de piedras muy menudas tales como silicatos y piedras areniscas y se utiliza en digerir las semillas sin pelar. Los pájaros, tales como palomas y gallinas, que tragan la semilla entera, requieren tal arena. La arena insoluble no se puede digerir y permanecerá en el cuerpo hasta excretarlo.

- La arena soluble es orgánica e incluye cáscaras machacadas (tales como cáscaras de ostra o de cascara de huevo esterilizada) o hueso de jibia. La arena soluble es sobre todo piedra caliza (carbonato de calcio). Puesto que es fácilmente digerido por los ácidos encontrados en el proventriculo( estómago glandular), hay poco peligro de que se acumule en el sistema digestivo. Sin embargo, mientras que la arena soluble es una fuente de calcio, también ayuda en la digestión de semillas enteras. Además, si no sabemos la procedencia de este grit podremos encntrarnos con casos de intoxicación por metales pesados los cuales proceden de arena soluble que ha venido de áreas de aguas contaminadas.

¿Qué pajaros necesitan grit(arena insoluble)?
Puesto que el propósito de la arena es quitar las capas externas de semillas enteras, solamente los pájaros que consumen las semillas enteras, requieren grit en su dieta.
La arena no se requiere en pájaros normales y sanos, psitácidas (loros) o paserines (canarios, pinzones). Puesto que estos pájaros quitan los cascos fibrosos de las semillas en el acto de comerlas, las enzimas digestivas no tienen ninguna barrera para evitar que actúen sobre la semilla. De hecho ciertas especies de loros tienen arrugas en la porción interior de su pico superior que se crean para ayudar a descascar las semillas. La semilla es sostenida por las arrugas del pico, mientras que el pico de abajo se utiliza para agrietar y para quitar la cáscara. Los pájaros con una dieta de pienso extrusionado(pellets) no requieren grit.
El libro AVIAN MEDICINE; Principles and Application nos dice: han habido muchos casos de pájaros sanos que no han tenido grit por períodos de 15 a 20 años y no han demuestrado ninguna señal de mala digestión.
Por otra parte, también se cree que pájaros con problemas pancreáticos o de la digestión físicos, pueden beneficiarse de algunos tipos de grit. Sin embargo, en tales casos debe consultarse a un veterinario aviar.
En los E.E.U.U., el uso del grit generalmente se desaconseja, especialmente si está ofrecido libremente ya que puede conducir a la gastritis obstructiva. En Australia, en donde el grit se da con frecuencia a los pájaros doméstico, se hallado pocos problemas. Por el momento que no hay explicación para estas diferencias geográficas.
Si un dueño siente que debe dar grit, el libro Avian Medicine recomienda que no deben ser ofrecidos libremente, sino como una dosificación ,a pájaros del tamaño del periquito se les pueden dar 5 granos de grit bianualmente (cada 2 años) y los pájaros como las cacatúas pueden darse media cucharilla cada 2 años.
Problemas potenciales causados por Grit:
Si se les da las aves acceso libre al grit, algunos pájaros pueden consumir más de lo debido, produciendo un grave impacto proventricular o ventricular.
Según Avian Medicine: “este problema se ha hallado comúnmente en Norteamérica pero aparece infrecuentemente en Australia. La causa de la variación regional en la ocurrencia de esta condición es desconocida. Los pájaros que demuestran la consumición obligatoria de grit deben ser evaluados de enfermedades hepáticas, pancreatitis, disfunción renal y desnutrición general. “
Es también importante comprobar el contenido de cualquier mezcla del grit comprada para saber si hay la presencia del carbón de leña. El carbón de leña puede afectar la absorción de las vitaminas A, B2 y K, dando por resultado deficiencias.
En relación con dietas formuladas( pienso extrusionado) Avian Medicine nos dice: Los “estudios en aves de corral indican que la adición de grit aumenta la digestibilidad en mas mas de un 10%, pero estudios similares no se han realizado en pájaros de compañía. Dado que la obesidad es un problema mayor que la mala digestión en pájaros de compañía, aumentando la digestibilidad de la dieta formulada se sugirió que los requisitos alimenticios del grit son probablemente innecesarios.
En conclusión, los beneficios del grit para los loros no se ha demostrado, pero si se han observado los riesgos potenciales
Fuentes;
Avian Medicine: Principles and Application by Ritchie, Harrison and Harrison.
http://www.birdsnways.com/wisdom/ww58eii.htmhttp://www.bellaonline.com/articles/art1529.asp
Aurora Carranza Román
sábado, noviembre 25, 2006
EL MITO DE LA MONOGAMIA EN AGAPORNIS
Mucha gente cree en el mito de que los agapornis son aves inseparables y que si las separamos de su pareja o esta muere caerán en una depresión muriendo ellas también,pues bien todo esto es exagerado, bien es cierto que son aves sociables y que elaboran vinculos de unión fuertes entre ellos, algunos, sobre todo los que estan separados en jaulas y no tiene contacto alguno con seres humanos o otras especies, al morir su pareja si que sufren, pero porque son seres muy sociables.

El agaporni es tan promiscuo como otras muchas especies pequeñas de loro(periquitos, forpus.....). Es un mito que estos pájaros forman enlaces para siempre. De hecho si lo pensais transladandolo a su estado salvaje, esto seria un callejón sin salida evolutivo para los agapornis ya que la supervivencia de la especie es dependiente de las uniones e intercambios de genes. Ésto no quiere decir que los agapornis no sigan emparejados o juntos para toda la vida vida, pero las relaciones monogamas son amenudo definidas como una union con una pareja para toda vida y el concepto de unión de estas aves no es el mismo. Si lo piensas bien la monogamia no sería beneficiosa en su estado salvaje, donde la muerte de una de pareja requiere el reemplazamiento de su pareja inmediato por otro animal, las hembras y los machos poligamos o promiscuos son deseables en la situación de colonia, pues demuestra su buena salud reproductiva, aunque ésta no se traduce en buenos padres ni que sean buenos padres adoptivos aunque siempre hay excepciones.
La estabilidad de la pareja mejora tras criar juntos una nidada o dos, pero no es una garantía de monogamia en una colonia, en mi caso despues de criar con varias parejas en pajarera y haber obtenido 3 nidadas de cada uno, tras cambiarlos de sitio se intercambiaron las parejas, fastidiandome los cruces, por lo que tuve que volver a obligarlas a criar como queria.
De hecho, en una colonia si te fijas veras a menudo parejas estables que permanecen juntas, pero en algun momento mientras el macho se descuida la hembra recibe caricias y las atenciones de otros machos hasta dejandose copular por otros a veces, mientras que el macho tambien puede visitar a otras hembras, mientras su pareja oficial incuba los huevos de otro macho/s o de el mismo.Este patron no se observa en emparejamientos por parejas en jaula y de hecho es lo que los criadores exactamente evitamos que suceda emparejandolos en jaulas.
La hembra a veces rechazará copular con el macho después de dos nidadas con él, pero él continuará alimentandola y ocupándose de la hembra y de los pichones , esta forma de emparejamiento se le denomino en el pasado patron social monogámico , pero no es asi pues mantiene enlaces con su pareja pero practica la infidelidad.
Esto explicaría un poco lo de las parejas del mismo sexo que vemos en una colonia si ocurre una representación desigual de sexos (mas hembras que machos y al contrario)se emprejan y muestran signos de sociabiliad entre ellos ,pero llegado el momento aceptan emparejarse y copular con el sexo opuesto, estas parejas del mismo sexo puede que no se separen después de varios años de interacción, y algunas son buenas incubadoras y alimentan bien a sus pollos mientras que mantenienen lazos con el mismo sexo (hembra- hembra pero en macho-macho no es tan seguro ).
El agaporni es tan promiscuo como otras muchas especies pequeñas de loro(periquitos, forpus.....). Es un mito que estos pájaros forman enlaces para siempre. De hecho si lo pensais transladandolo a su estado salvaje, esto seria un callejón sin salida evolutivo para los agapornis ya que la supervivencia de la especie es dependiente de las uniones e intercambios de genes. Ésto no quiere decir que los agapornis no sigan emparejados o juntos para toda la vida vida, pero las relaciones monogamas son amenudo definidas como una union con una pareja para toda vida y el concepto de unión de estas aves no es el mismo. Si lo piensas bien la monogamia no sería beneficiosa en su estado salvaje, donde la muerte de una de pareja requiere el reemplazamiento de su pareja inmediato por otro animal, las hembras y los machos poligamos o promiscuos son deseables en la situación de colonia, pues demuestra su buena salud reproductiva, aunque ésta no se traduce en buenos padres ni que sean buenos padres adoptivos aunque siempre hay excepciones.
La estabilidad de la pareja mejora tras criar juntos una nidada o dos, pero no es una garantía de monogamia en una colonia, en mi caso despues de criar con varias parejas en pajarera y haber obtenido 3 nidadas de cada uno, tras cambiarlos de sitio se intercambiaron las parejas, fastidiandome los cruces, por lo que tuve que volver a obligarlas a criar como queria.
De hecho, en una colonia si te fijas veras a menudo parejas estables que permanecen juntas, pero en algun momento mientras el macho se descuida la hembra recibe caricias y las atenciones de otros machos hasta dejandose copular por otros a veces, mientras que el macho tambien puede visitar a otras hembras, mientras su pareja oficial incuba los huevos de otro macho/s o de el mismo.Este patron no se observa en emparejamientos por parejas en jaula y de hecho es lo que los criadores exactamente evitamos que suceda emparejandolos en jaulas.
La hembra a veces rechazará copular con el macho después de dos nidadas con él, pero él continuará alimentandola y ocupándose de la hembra y de los pichones , esta forma de emparejamiento se le denomino en el pasado patron social monogámico , pero no es asi pues mantiene enlaces con su pareja pero practica la infidelidad.
Esto explicaría un poco lo de las parejas del mismo sexo que vemos en una colonia si ocurre una representación desigual de sexos (mas hembras que machos y al contrario)se emprejan y muestran signos de sociabiliad entre ellos ,pero llegado el momento aceptan emparejarse y copular con el sexo opuesto, estas parejas del mismo sexo puede que no se separen después de varios años de interacción, y algunas son buenas incubadoras y alimentan bien a sus pollos mientras que mantenienen lazos con el mismo sexo (hembra- hembra pero en macho-macho no es tan seguro ).
Aurora Carranza Román
jueves, noviembre 23, 2006
BIODIVERSIDAD Y ESTADO ACTUAL DE LOS AGAPORNIS
BIODIVERSIDAD
En Biología el término biodiversidad abarca todo lo relativo a la diversidad de la vida, tanto en lo que se refiere a la diversidad de las formas de vida como en lo referido a la diversidad y complejidad interna de los ecosistemas. El Día Internacional se celebra el 22 de mayo.
La biodiversidad existente en la Tierra actualmente es el resultado de más de 4 mil millones años de evolución
Por causas naturales, la Tierra ha conocido momentos de extinción masiva de especies a lo largo de su historia geológica, que han reducido drásticamente la biodiversidad.
Después de cada gran extinción, las oportunidades representadas por los nichos que quedan vacantes favorecen nuevas radiaciones evolutivas y, en conjunto, la nueva diversificación que sigue a una extinción consigue al cabo de unas decenas de millones de años superar la diversidad anterior.
Se considera generalmente que la expansión demográfica y económica de la especie humana está poniendo en marcha una extinción masiva, de dimensiones incomparablemente mayores que las de cualquier extinción anterior.
Las causas concretas están en la desaparición indiscriminada de ecosistemas, por la tala de bosques, la degradación de los suelos, la contaminación ambiental, la caza y la pesca excesivas, ...etc. La comunidad científica juzga, en general, que tal extinción representa una amenaza para la capacidad de la biosfera para sustentar la vida humana a través de diversos servicios naturales y recursos renovables.
La biodiversidad ha contribuido de muchas maneras al desarrollo de la cultura humana, y, a su vez, las comunidades humanas han jugado un papel mayor formando la diversidad de naturaleza al genético, especies y los niveles ecológicos.
IUCN (UNION INTERNACIONAL PARA LA CONSERVACION DE LA NATURALEZA Y LOS RECURSOS NATURALES)
La Unión Internacional para la Conservación de la Naturaleza y los Recursos Naturales (International Union for Conservation of Nature and Natural Resources, abreviado IUCN) es un organismo internacional dedicado a la conservación de los recursos naturales.
Fundada en 1948, tiene su sede en Gland, Suiza. La IUCN reúne a 78 estados, 112 agencias gubernamentales, 735 ONGs y miles de expertos y científicos de 181 países.
La misión de la IUCN es la promoción y asistencia a las sociedades en todo el mundo para que conserven la integridad y biodiversidad de la naturaleza y aseguren que el uso de los recursos naturales es equitable y ecológicamente sustentable.
La pérdida de la diversidad biológica es una de las crisis más apremiantes del mundo y la preocupación sobre el estado de los recursos biológicos de los cuales depende significativamente la vida humana está aumentando. Se ha estimado que la tasa de extinción actual es de 1 000 a 10 000 veces superior a la que ocurriría naturalmente.
Muchas especies están disminuyendo a niveles poblacionales críticos, hábitats importantes han sido destruidos, fragmentados y degradados, y los ecosistemas han sido desestabilizados a través del cambio climático, la polución, las especies invasoras y el impacto humano directo. Sin embargo, la mayor conciencia sobre la manera en que la diversidad biológica sustenta los medios de subsistencia, permite el desarrollo sostenible y la cooperación alentadora entre naciones. Esta conciencia es generada a través de productos como la Lista Roja de la UICN.
Lista roja
La Lista Roja de la IUCN es una lista de especies, subespecies, variedades o incluso poblaciones de seres vivos amenazados, en diferentes grados, de extinción, fundada en 1965 a nivel mundial.
Existen nueve categorías en el sistema de la Lista Roja de la UICN:
- Extinto,
- Extinto en Estado Silvestre
- En Peligro Crítico
- En Peligro
- Vulnerable
- Casi Amenazado,
- Preocupación Menor
- Datos Insuficientes
- No Evaluado.
(EX) EXTINTO ; Un taxón está Extinto cuando no queda ninguna duda razonable de que el último individuo existente ha muerto. Se presume que un taxón esta Extinto cuando prospecciones exhaustivas de sus hábitats, conocidos y/o esperados, en los momentos apropiados (diarios, estacionales, anuales), y a lo largo de su área de distribución histórica, no ha podido detectar un solo individuo. Las prospecciones deberán ser realizados en períodos de tiempo apropiados al ciclo de vida y formas de vida del taxón.
(EW) EXTINTO EN ESTADO SILVESTRE ; Un taxón está Extinto en Estado Silvestre cuando sólo sobrevive en cultivo, en cautividad o como población (o poblaciones) naturalizadas completamente fuera de su distribución original. Se presume que un taxón está Extinto en Estado Silvestre cuando prospecciones exhaustivas de sus hábitats, conocidos y/o esperados, en los momentos apropiados (diarios, estacionales, anuales), y a lo largo de su área de distribución histórica, no han podido detectar un solo individuo. Las prospecciones deberán ser realizadas en períodos de tiempo apropiados al ciclo de vida y formas de vida del taxón.
(CR) EN PELIGRO CRITICO; Un taxón esta En Peligro Crítico cuando la mejor evidencia disponible indica que cumple cualquiera de los criterios "A" a "E" para En Peligro Crítico y, por consiguiente, se considera que se está enfrentando a un riesgo extremadamente alto de extinción en estado silvestre.
(EN) EN PELIGRO Un taxón esta En Peligro cuando la mejor evidencia disponible indica que cumple cualquiera de los criterios "A" a "E" para En y, por consiguiente, se considera que se está enfrentando a un riesgo muy alto de extinción en estado silvestre.
(VU) VULNERABLE Un taxón es Vulnerable cuando la mejor evidencia disponible indica que cumple cualquiera de los criterios "A" a "E" para Vulnerable y, por consiguiente, se considera que se está enfrentando a un riesgo alto de extinción en estado silvestre.
(NT) CASI AMENAZADO Un taxón está Casi Amenazado cuando ha sido evaluado según los criterios y no satisface, actualmente, los criterios para En Peligro Crítico, En Peligro o Vulnerable; pero está próximo a satisfacer los criterios, o posiblemente los satisfaga, en el futuro cercano.
(LC) PREOCUPACIÓN MENOR Un taxón se considera de Preocupación Menor cuando, habiendo sido evaluado, no cumple ninguno de los criterios que definen las categorías de En Peligro Crítico, En Peligro, Vulnerable o Casi Amenazado. Se incluyen en esta categoría taxones abundantes y de amplia distribución.
(DD) DATOS INSUFICIENTES Un taxón se incluye en la categoría de Datos Insuficientes cuando no hay información adecuada para hacer una evaluación, directa o indirecta, de su riesgo de extinción basándose en la distribución y/o condición de la población.
(NE) NO EVALUADO Un taxón se considera No Evaluado cuando todavía no ha sido clasificado en relación a estos criterios.
Una vez explicados estos términos vamos a extrapolarlos a los agapornis y a valorar la situación actual de estos.Para empezar voy a hacer un breve recordatorio de la clasificación cientifica de nuestros pájaros y de cómo estan emparentados siguiendo la evolución de la especie.
La Filogenia es la disciplina que estudia las relaciones evolutivas entre las distintas especies, reconstruyendo la historia de su diversificación (filogénesis) desde el origen de la vida en la Tierra hasta la actualidad. La filogenia proporciona el fundamento para la clasificación de los organismos, según esto tenemos esta clasificación filogénica de los agapornis;

Reino; Animal
Filo; Chordata
Clase; Aves
Orden ; Psittaciformes
Familia; Psittacidae
Género; Agapornis Selby, 1836
Para llegar a esta clasificación y a este árbol filogénico, se han hecho estudios del comportamiento de los agapornis y estudios genéticos de ellos, utilizandose la prueba del mtADN, que continuación explicare un poco lo qué es;
Las dos más comunes pruebas de genealogia ADN son la de cromosoma Y o Y-DNA (linea paterna) y la de mitocondria o mtDNA (linea materna). El objetivo de estas pruebas es el comparar a una especie de agaporni con otra para determinar la probabilidad numerica que los dos individuos esten emparentados dentro de cierto numero de generaciones.
Linaje paterno
El ADN de la cromosoma Y o ADN-Y (Y-DNA en ingles) se pasa de una generacion a la siguiente solo de varon a varon. Como los apellidos tambien pasan de una generacion a la siguiente de varon a varon (en las culturas de origen no hispano), esta prueba es la mas adecuada para comparar individuos con el mismo apellido, o por lo menos con sospecha de parentezco.
Linaje materno
El ADN mitocondrial o ADN-mt (mtDNA en ingles) se pasa de una hembra a todos sus hijos e hijas, tanto machos como hembras. Sin embargo, el ADN-mt solo pasa a la siguiente generacion por las hembras. Por tanto solo hay un pequeño numero agapornis en su arbol familar que comparten su ADN-mt. Todos estos agapornis son descendientes por la rama femenina de su ancestro femenino mas antiguo. Sus hijos e hijas tambien comparten su ADN-mt, pero solo si es una hembra. Sus descendientes machos no pueden pasar su ADN-mt a sus descendientes.
Según el estudio de la prueba mtADN se demuestran que;
- Compartieron todos un antepasado común relativamente reciente
- Existe una relación cercana entre el taranta y el pullaria.
- Existe una relación levemente más cercana entre los nigrigenis y los lilianae.
Segun Moreau , la separación entre los clanes nigrigenis/lilianae y personata/fischeri probablemente fue causada por las montañas en la cabecera del lago Nyasa.
Al norte la división de personata/fischeri coincide con la localización del Rift Valley, que puede ser responsable de su separación; sin embargo, en el final meridional de la división, no hay barreras geográficas importantes que separen a estas dos especies (Moreau 1948).Moreau (1948) sugirió que la separación se produjo por las regiones del bosque "miyombo" y, en el caso de nigrigenis y de lilianae, por una región de la alta elevación montañosa.
Ahora pasaremos a ver el mapa de distribución de las especies.
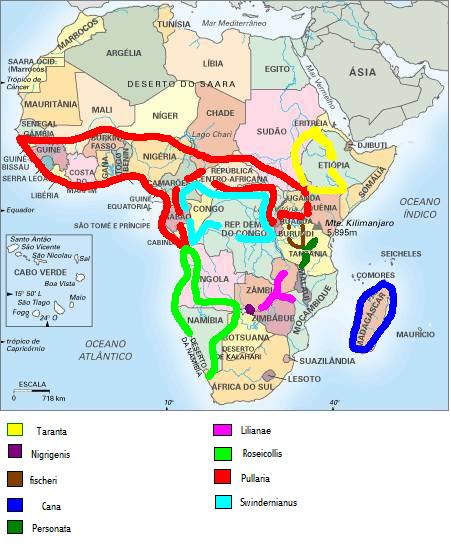
Una vez repasada esta información vamos a volver al tema de la biodiversidad y de la importancia de conservar la especie pura en todos los agapornis, ya que algunos ya se estan viendo amenzados a pasos agigantados y quién sabe si en un futuro las demas especies no corran la misma suerte, por eso desde aquí quiero recordaros que estoy a favor de las mutaciones siempre y cuando se respete la forma ancestral o salvaje.
Debemos seguir criando la especie pura ya que si alguna catástrofe ocurriera, la repoblación estaría en manos de los humanos, por eso debemos estudiar a todas las especies e intentar criar en cautividad las menos proliferativas, para mantener la biodiversidad de los agapornis.
Estado actual de la lista roja de los agapornis
Fue actualizada por ultima vez en el 2004.
Los términos siguientes y su significado;
Threatened; Amenazado
Least concern; poca preocupación
Extint; extinto
Agapornis Roseicolli ...................preocupación menor

Agapornis Personata...................preocupación menor

Agapornis Cana..........................preocupación menor

Agapornis Pullaria......................preocupación menor

Agapornis Taranta......................preocupación menor

Agapornis Swinderniana.............preocupación menor

Agapornis Fischer…………………casi amenzado

Agapornis Lilianae.............casi amenazado

El Agapornis Lilianae es originario del sur de Tanzania, norte de Zimbabwe y el este de Zambia, habita actualmente en el Liwonde Nacional Park (LNP) y algunos grupos viven en los bosques circundantes del LNP .
Su distribución rápidamente se está restringiendo a LNP porque sus hábitos de alimentación y cría se están alterando debido a la explotación agrícola.
Este parque se esta viendo afectado por el crecimiento y el desarrollo de la población. Recientemente, los casos del envenenamiento de Agapornis Nyasa se han intensificado aunque no se sabe porqué los cazadores furtivos están envenenando los pájaros.
Agapornis Nigrigenis .........................vulnerable

Esta especie es vulnerable puesto que tiene una población pequeña la cual esta experimentando una baja continua debido a pérdida y a la degradación del hábitat, particularmente por la desecación gradual de los sitios donde van a beber agua.
Desde 1950, la precipitación anual en el hábitat de los nigrigenis ha disminuido, aumentando la dependencia de abastecimientos de agua artificiales. En este estudio (Conservation implications of the drinking habits of Black-cheeked Lovebirds Agapornis nigrigenis in Zambia), se observaron e investigaron los requisitos estacionales del agua y el comportamiento de beber de los agapornis en su hábitat natural.
Durante la estación seca, los agapornis nigrigenis bebieron en el mismo punto de agua por la mañana temprano y al final de la tarde. Las cantidades de la multitud de pájaros en los sitios que bebían se extendieron de 1 a 175 individuos. Los agapornis nigrigenis vigilaban y eran altamente cautelosos y no bebieron en los abastecimientos artificiales cuando eran molestados por los seres humanos o el ganado.
Las implicaciones para la conservación de la especie se discuten, ya que la disponibilidad del agua es un factor limitador para el nigrigenis y la desecación gradual de su habitat.
Debido a la dependencia de estas aves por el agua, se debe realizar un aumento de abastecimientos de agua artificiales, la prioridad para la gerencia de la conservación de la especie debe ser crear y mantiener recursos de agua evitando molestar a las aves para que no se asusten.
También hay otra teoria que se une a la anterior, la cual apunta a un aumento de una afección virica en esta especie(posiblemente BFDV), la cual ha derivado en un descenso de la población por muerte de individuos adultos y embriones y pollos.
Por último aqui os muestro un enlace de fotos de agapornis en su estado salvaje.
http://www.freewebs.com/personatus/sfeerfotos.htm
Fuente y artículos relacionados;
http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=412457
http://www.ajol.info/viewarticle.php?id=22968
http://www.ajol.info/viewarticle.php?id=22962
http://www.publish.csiro.au/nid/96/paper/MU04037.htm
Articlulo sobre el BFDV encontrado en agapornis en su habitat natural;
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=506959 http://www.findarticles.com/p/articles/mi_qa3793/is_199804/ai_n8785457/pg_6 http://www.ingentaconnect.com/content/nisc/ostrich/2005/00000076/F0020003/art00009 http://www.findarticles.com/p/articles/mi_qa3793/is_199804/ai_n8785457
IUCN 2004. 2004 IUCN Red List of Threatened Species. <http://www.iucnredlist.org/>. Downloaded on 14 December 2004.
http://elibrary.unm.edu/sora/Auk/v115n02/p0455-p0464.pdf
Aurora Carranza Román
sábado, noviembre 11, 2006
HIBRIDACIÓN Y TRANSMUTACIÓN
LA HIBRIDACIÓN Y LA TRANSMUTACIÓN
Un tema muy comentado y que suscita muchas preguntas y pólemica es el tema de la hibridación, para ello expondre varias de las preguntas frecuentes y su respuestas, espero que os sean de ayuda.Primero haremos un esquema de las especies con anillo y sin anillo ocular ( también llamado perioftálmico).
Con anillo ocular tenemos;
-Agapornis personata.Fue presentado a la ciencia por Reichenow en 1887.
-Agapornis fischer.Fue presentado por Reichenow en 1887.
-Agapornis lilianae.Fue presentado a la ciencia por Shelley en 1894
-Agapornis nigrigenis.Presentado por Sclater, WL, en 1906.
Sin anillo ocular;
-Agapornis cana. Tenemos dos subespecies; agapornis cana cana, presentado a la ciencia por Gmelin en 1788, y agapornis cana ablectanea por Bangs en 1918
-Agapornis roseicollis.Tenemos dos subespecies, roseicollis roseicollis, presentado a la ciencia por Vielliont 1917 y roseicollis catumbella por Hall en 1952
-Agapornis taranta.Tenemos dos subespecies, taranta taranta presentado a la ciencia por Stanley en 1814, y taranta nana por Neumann en 1931
-Agapornis swinderniana.Tenemos 3 subespecies, agapornis swindernianus swindernianus( Kuhl, 1820), agapornis swindernianus emini ( Neumann, 1908) y agapornis swindernianus zenkeri ( Reichenow,1895)
- Agapornis pullaria.Tenemos 3 subespecies, agapornis pullarius pullarius (Linnaeus, 1758), agapornis pullarius guineensis ( Statius Müller, 1776), agapornis pullarius ugandae (Neumann, 1908).
Mucha gente hibrida por desconocimiento, porque les venden en las tiendas pájaros que ni ellos mismo saben qué son, por informaciones erróneas y por estar mal aconsejados. Por lo que debemos estar informados si queremos tomarnos en serio esta afición y queremos disfrutar de la belleza de los agapornis, ya sabeis nadie nace sabiendo, por lo que lo mejor es preguntar mil veces lo que no se sepa y leer mucho y aprender de los criadores mas experimentados y que de verdad sepan de lo que estan hablando.
¿Qué especies al cruzarlas dan descendencia fértil?
Si cruzamos especies de anillo ocular( lilianae, nigrigenis, personata y fischer) estas crias serán fértiles y no seran ni una cosa ni otra, estos pájaros al cruzarlos nuevamente daran híbridos y todos los cruces posteriores segurian dando ejemplares fértiles.
Hay también gente que habla de cruces de taranta con roseicollis los cuales carecen de anillo y tambien serían fértiles, pero yo como no tengo constancia segura de ello no lo desarrollare más.
¿Qué especies al cruzarlas dan descendencia infértil?
Cuando cruzamos especies de anillo acular con no ocular(roseicollis, tarantas..)la descendencia será infértil y estos pájaros no podran tener descendencia, esto no quiere decir que no tengan instinto reproductor, ya que incluso se aparean y ponen huevos como los demás, pero los huevos son infértiles, ya los crucemos con otros infértiles o con ejemplares puros no habra fecundación, y por no tanto no habra embrión.
Mención aparte merecen los comentarios de que estos cruces no generan agapornis estériles en su totalidad, y que estos hibridos necesitan más tiempo e incluso varios años para madurar y criar, yo personalamente no tengo constancia visual de ello, se comenta que es cierto, pero en todo caso no hagais la prueba.
Después de estas explicaciones quiero decir desde aquí que no debemos cruzar especies, ya que algunas de ellas son dificiles de encontrar puras y algunas de ellas son escasas en su hábitat natural.
¿Qué es una transmutación?
Cuando se quiere pasar un color determinado de una mutación de una especie a otra, hablamos de transmutación, y se hace entre ejemplares que dan descendencia fértil evidentemente, como ejemplo, la mutación lutino surgio por primera vez en los agapornis lilianae en Australia sobre el 1936, y como esta mutación no existía en las demás especies se cruzaron ejemplares lutinos lilianae con personatas para pasar esa mutación a los personata, y de los personata se paso posteriormente a los fischer. Lo mismo paso con la mutación azul que se paso a través de transmutación de personata a fischer.
Estas transmutaciones deben hacerlas criadores expertos y no cualquiera que quiera experimentar.Por lo tanto transmutar es hibridar entre ejemplares que dan descendencia fértil de forma controlada y por expertos en la materia.
Para trasmutar se debe tener una excelente línea pura para que cuando hagamos la transmutación sucesivamente se vayan emparejando estos ejemplares que son hibridos e ir acercándolos hacia el standard o color puro, por lo que este proceso conlleva muchos años de selección y lo peor de todo son esos ejemplares intermedio(hibridos) que quedan disponibles.
Os preguntareis qué ocurre con esos pajaros, hay dos versiones, sacrificar a todos esos pajaros intermedios para asi no dañar las líneas de las especies puras o lo que yo creo que se suele hacer, que es venderlos como puros llenando el mercado de hibridos.
Un criador honesto que transmuta debera informar a sus compradores de lo que realmente cria,pero no siempre es así, tambien se da el caso de criadores que realmente no saben que sus pajaros no son puros ya que algunas mutaciones son dificiles de distinguir, y los venden sin mala intención,pero perjudicando a la especie y haciendo mas difícil la tarea de conseguir ejemplares puros.
Bajo mi punto de vista sacrificar a las crias para conseguir un tipo de mutación a base de manipular y matar aves no lo veo correcto,pero se hace y todos disfrutamos de estos colores sin pensar en ello, ahi cada cual tiene su criterio y no profundizare en él.Aunque como ya he dicho anteriormente yo creo que la mayoria de estos hibrídos los venden a importadores y pajarerias.
Cuando una mutación se pasa de una especie a otra hasta la sexta generación no podemos decir que tenemos pájaros puros de esta mutación, cuando el porcentaje de la pureza es superior al 98%, Ahora hare un pequeño esquema de una trasmutación:
- fischer puro x personata puro = personata-fischer (50% sangre fischer y 50% sangre personata).F1
- fischer puro x F1 = personata-fischer ( 75% sangre fischer y 25% sangre personata )F2
- fischer puro x F2 = personata-fischer ( 87,5% sangre fischer and 12,5% sangre personata)F3
- fischer puro x F3 = personata-fischer ( 93,75% sangre fischer y 6,25 % sangre personata)F4
- fischer puro x F4 = personata-fischer ( 96,87% sangre fischer y 3,13% sangre personata)F5
- fischer puro x F5 = personata-fischer ( 98,43% de sangre fischer y 1,57% sangre personata) F6
Y para alcanzar estos porcentajes de pureza de sangre tenemos que estar seguros que el fischer que cruzamos con el primer personata y con el persofischeri (F1, el F2…)son puros y ya sabemos lo dificil que es esto.
Los F son ejemplares intermedios( hibridos) y conforme los cruzamos con puros vamos adquiriendo pajaros mutados con más porcentaje de sangre fischer , asi se hacen cruces sucesivamente hasta la quinta o sexta generación que es en la cual se supone que obtenemos ejemplares con sangre fischer por lo menos superiores al 98%, por lo que los pajaros transmutados mantienen algunos signos minimos de sangre personata( por ese 1,57%) ,que se podran mostrar en su descendencia y los cuales debemos trabajar en nuestros pájaros.
Para mi un pájaro mutado puro es un ave que siendo mutada mantiene caracteristicas similares con el ancestral, como ejemplo los fischer azules, deben de tener la eumelanina de la cabeza igual que un ancestral bueno ya que lo que se elimina es la psitacina y la eumelanina debe queda intacta, y no tener la cara mancha con intromisión del negro, ni una linea horizontal por detrás de la nuca signo de la trasmutación con personata, deben de tener una linea diagonal por detrás de la cabeza que es lo que se exige en concursos.
Si queremos hacer la prueba podemos cruzar nuestros pájaros mutados que consideremos buenos con un ancestral( verde)bueno, si la descendencia se aleja mucho del ancestral entonces nuestros pájaros no son todo lo puros que pensabamos y se debería trabajar mas esa línea, cruzándolos con verdes puros, tarea dificil y que lleva años de dedicación, si por el contrario la descendencia es como uno puro ancestral, tendremos pajaros buenos y con esos se trabajara mejor.
Por último desde aqui quiero decirle a la gente que este leyendo mi artículo que no hibride, que deje las transmutaciones para gente experta, ya que el resultado casero de hibridar no nos lleva a ningún lado ni supone un gran mérito para el criador aficionado ya que es fácil conseguirlo(hibridar entre dos especies), y mediante esto no vamos a descubrir mutaciones nuevas, solo pájaros intermedios que en el caso de ser fértiles, nos haran mas difícil la tarea de conseguir pájaros puros y que ensucian nuestra afición.
Aurora Carranza Román
sábado, noviembre 04, 2006
MUTACIONES DE COLOR EN LOS AGAPORNIS
MUTACIONES DEL COLOR EN LOS AGAPORNIS
El color ancestral es color salvaje que tienen los agapornis en su hábitat.Cada especie de agapornis tiene un color característico.
Ahora para entender mejor las mutaciones repasaremos algo de genética y de la luz.
Los cromosomas van de dos en dos, en parejas, y los genes estan en los cromosomas por lo que tambien van de dos en dos.
Nos encontramos con 2 tipos de mutaciones;
1. Dominantes; Son las mutaciones en las cuales con las que solo este presente en uno de los genes( es lo mismo que en uno de los cromosomas) se muestran fenotipicamente. estas a su vez se subdividen en ;


Dibujo 1
- Dominantes completas; en las que no hay diferencia fenotípicas ( de color) entre los SF (factor simple) y DF ( factor doble)
- Dominates incompletas o codominantes; hay diferencias de color entre un SF y un DF
Las abreviaturas SF y DF van detrás de la mutación dominante (nunca de una recesiva) significan;
- SF; factor simple,


- DF; factor doble, cuando el gen dominate esta en ambos cromosomas, ejemplo en este dibujo;


2. Recesivas; Son las mutaciones en las que se tiene que dar los dos genes, en los dos cromosomas para que la mutación se vea .
Si solo hay un gen recesivo;
El pájaro sea fenotipicamente verde y portador por lo tanto de una mutación recesiva
Alelos multiples
Los alelos múltiples son formas diferentes, expresiones del mismo gen es decir versiones diferentes de un gen
Del mismo modo, algunos genes del color se manifiestan en diferentes versiones.
Intentare explicar toda esta terminologia con unos dibujos
Los cromosomas van de dos en dos, en parejas, y los genes estan en los cromosomas por lo que tambien van de dos en dos.
Para que una mutación recesiva se vea es necesario que este en los dos cromosomas;


C1 C2
En este dibujo vemos C1(cromosoma 1) y C2( cromosoma 2), vereis que en uno de los brazos de C1 hay un hueco color amarillo y en C2 un hueco color azul, pues bien ese hueco representa el locus(lugar que ocupa un gen) y el color representa a los genes, el amarillo sera INO y el azul DEC, estos dos genes son recesivos, por lo que tendrian que darse en los dos cromosomas para verse, pero como son genes que ocupan el mismo lugar en el cromosoma cada uno va a ocupar un hueco dandose un color intermedio
El locus (NSL ino) puede también mutarse en pastel y en dark eyed clear. Por lo que;
- un gen codificado para ino, es un alelo para pastel
- un gen codificado para ino, es un alelo para dark eyed clear
- un gen codificado para pastel, es un alelo para dark eyed clear.
Estos genes emparentados se influencian mutuamente por lo que cuando los encontramos juntos en un mismo pájaro aparece un fenotipo intermedio.
Si el pastel reduce la melanina un 50% y el ino un 100%. La combinación de pastel e ino lo hace un 75%.
Este color no podemos llamarlo pastel/ino (pastel portador de ino).porque implicaría la dominancia del pastel sobre el ino y no habría diferencia visual entre por ejemplo, un verde pastel y un pastel verde/ino.
Por lo tanto no existen ;
- pastel/ino, dec/ino, pastel/dec, esto quiere decir que nunca un pájaro lutino podra portar ni dec ni pastel, un pájaro dec no podra portar ino ni pastel, un pájaro pastel no podra portar ino ni dec.
Tampoco un pastelino, decino, pasteldec puede portar una tercera de estas 3 que le falte, ya que cada cromosoma tiene un hueco y solo hay dos espacios.
Los nombres que debemos utilizar para estos fenotipos intermedios son pastelIno, decIno y pastelDec y estas mutaciones se escriben con una palabra, ni separandolas con espacios, ni guiones.
En Agapornis roseicollis se produce un fenómeno similar pero con los cromosomas autosómicos recesivos y ligados al sexo que son;
-Aqua y turquesa pueden combinarse en aquaturquesa ( cromosomas autosomicos recesivos)
- el recesivo ligado al sexo ; pallid e ino en pallidino( solo los machos pueden serlo ya que el cromosoma de las hembras Y(W) no codifica color)
Tampoco existen los;
-Aqua/turquesa, pallid/ino
REDUCCIONES EN LOS PIGMENTOS DE LA EUMELANINA Y PSITACINA
- azul.................................reducción de psitacina del 100%
- ino................................ reducción cuantitativa de melanina del 100%. Hay dos tipos de mutacion Ino. Ino NSL significa Non Sex Linked ino (ino no ligado al sexo) y ino SL significa ino ligado al sexo y se da solo en roseicollis.
- Dark eyed clear............. reducción de melanina del 90%. Esta mutación es un alelomorfo múltiple del gen ino NSL
- dilute...............................reducción cuantitativa de melanina del 80%.
- Pastel...........................reducción cuantitativa de melanina del 50%.La mutación pastel es un alelomorfo múltiple del genino autosómico recesivo ( del ino NSL)
-Arlequin dominante; reduccion de eumelanina y psitacina roja de la mascara aleatoria.
- Arlequin recesivo; ausencia casi total de melanina, tambien hay una reduccion de la psitacina roja en la mascara dando como resultado una mascara mas pequeña.
- Pallid(roseicollis) o cinammon australiano; reduccion de eumelanina del 60%. (nacen con ojos rojos y a los 8 dias aproximadamente se oscurecen)
- Cinnamon o canela americano( roseicollis); en esta mutacion no hay una reduccion de eumelanina pero la he metido aqui para hacer la comparativa con el pallid, en esta ocasion lo que ocurre es una alteracion de la eumelanina que en vez de ser negra es marron, en esta mutacion tambien nacen con los jos rojos y a los 8 aproximadamente se oscurecen.
ALTERACIONES EN LA ESTRUCTURA DE LA PLUMA
- Violeta:
El factor violeta cambia el diámetro de las vacuolas de la zona esponjosa y causa una distribución diferente de las melanina en las barbas de la pluma. En vez de ondas azules de la luz se reflejan las violetas. El violeta se ve mejor en un pájaro azul que posee un factor oscuro(azul D o cobalto) y con dos factores violetas(DF). Tambien puede darse este factor violeta en pájaros de linea verde, pero no son tan bonitos como los de la linea azul.
- Factor de oscuridad( dark factor); en la que hay una alteración del ancho de la zona esponjosa de la pluma, a mas reduccion de esa zona mas oscuridad, por esta razón no debemos cruzar dos agapornis con dos factores de oscuridad, ya que esta mutación altera la calidad de la pluma y muchos de estos pichones resultado del cruce anterior no podran volar( no quiere decir que sea asi en todos). Ejemplo; malva x malva
Despues de esta escueta explicación de algunas de las mutaciones, debo deciros que se pueden dar a la vez varias mutaciones, es decir podemos tener un agaporni con una mutacion recesiva y a la vez otra dominate, o dos recesivas o incluso mas de dos juntas, como por ejemplo;
- fischer slaty malva pastel edged.
- roseicolli aqua arlequin edged.
- fischer cobalto violeta arlequin edged.
Tambien hacer mención que hay mutaciones que por su reducción excesiva de eumelanina y psitacina no podran mostrar una mutación aunque lo sean; ejemplo ;
-lutino jade, lutino oliva.
Y algunas mutaciones en las que haya una reduccion de eumelanina se pueden enmascarar unas a otras; ejemplo;
-un fischer arlequín puede ser edged, pero puede ser un arlequin muy marcado que sea dificil disitinguir las dos mutaciones sin saber su prodecencia genealogica.
Fuentes;
http://www.cockatielworld.org/genetics.html
Aurora Carranza Román
EL COLOR DE LOS AGAPORNIS
LA LUZ Y EL COLOR DE LOS AGAPORNIS
La luz es una onda electromagnética capaz de ser percibida por el ojo humano y cuya frecuencia determina su color.
La luz visible forma parte de una estrecha franja que va desde longitudes de onda de 380 nm (violeta) hasta los 780 nm (rojo). Los colores del espectro se ordenan como en el arco iris, formando el llamado espectro visible; ondas de luz rojas, naranjas, amarillas, verdes, azules, añiles y violetas

Hay dos tipos de objetos visibles: aquellos que por sí mismos emiten luz( estrellas como el sol) y los que la reflejan. Ejemplos;
- Un cuerpo que es rojo solo refleja las ondas de luz rojas
- Un cuerpo que es naranja solo refleja las ondas de luz naranjas
Pero en lo dicho anteriormente hay dos excepciones:
- Un cuerpo negro absorbe todas las ondas de luz
- Un cuerpo blanco las reflejas todas
Ahora vamos a traspasar estos conceptos a las plumas de nuestros agapornis :
Las barbas de la pluma y en menor grado las bárbulas combinadas con los pigmentos producen el color de los agapornis.
Las mutaciones de color se dan por ;
- cambios en la estructura de la pluma ( ejemplo; dark factor y violeta)
- por cambios en los pigmentos ( psitácina y eumelanina)
Del corte de una barba coloreada permite distinguir tres partes:
-El anillo extremo, llamado córtex, que contiene la psitacina
-La zona esponjosa, donde tiene lugar la difiracción de la luz solar.
-El centro, compuesto de la medula. La medula esta compuesta por innumerables células, en cuyo centro se halla una ampolla vacía o vacúola. Alrededor de cada vacúola se fijan los melanosomas
La luz natural incide sobre la barba de la pluma se difracta en la zona esponjosa, que contiene unas burbujas de aire que cumplen la misma función que las gotitas de agua que dan origen al arco iris y se divide en los siete colores del espectro.
Las ondas de luz amarilla son reflejadas por la psitacina amarilla en el córtex.
Las ondas de luz restantes penetran en la barba y en la zona esponjosa se forma una cantidad de luz azul y violeta por medio interferencias. Esta luz azul y violeta es enviada de nuevo por las vacuolas en la médula, mientras los otros colores son absorbidos por la melanina negra en la médula.
Así que finalmente tenemos una mezcla de ondas de luz amarillas y azules que se manifiestan como verdes, aunque sus plumas no contienen ningún pigmento ni colorante verde.
Si el amarillo externo (el córtex) no contiene colorante amarillo, los rayos azules llegarán a la retina sin atravesar la zona amarilla, permitiéndonos percibir entonces una coloración azul ( es lo que ocurre en la mutación azul en la que hay una reduccion del 100% de psitácida, que es el pigmento amarillo)
Cuando no hay una base negra( no hay melanina negra en la médula) la luz azul es invisible ya que no se absorben los colores si no que se reflejan. Y todos los colores combinados crean la luz blanca.
Los tonos como el azul y el violeta son originados por la refracción de los rayos de luz sobre el plumaje y su efecto varia según el ángulo de difusión de la luz. .
En forma resumida y como ejemplo;
- Color ancestral ;
(luz azul en una base de eumelanina) + luz amarilla = verde
(azul + negro) + amarillo = verde
- Mutacion con reduccion de la eumelanina(negra)
(ausencia de luz azul + ausencia de negro) + amarillo = amarillo
(transparente + transparente) + amarillo = amarillo
- Mutación con reduccion de pigmento psitácida (amarillo- rojo del cortex)
(azul + negro) + sin color = azul
- Mutación que elimina psitácida y eumelanina ;
(ausencia de azul + ausencia de negro) + sin color = blanco
(sin color) + sin color = blanco.

Fuentes;
Aurora Carranza Román
MUTACIONES DE LA EUMELANINA
Las mutaciones de la eumelanina se pueden dividir en 3:
-Reducciones cualitativas( calidad) ; es el albinismo son los Ino SL, Ino NSL, fallow y canela.
- En los ino NSL se anula casi la actividad de la tirosinasa por lo que no hay sintesis de pigmentos y se producen matrices incoloras, pero estas matrices aunque esten vacias se depositan en las plumas
- En inos SL; la actividad de la tirosinasa no se disminuye, pero como se han producido matrices pequeñas y deformadas la actividad de la tirosinasa no tiene efecto.
En los dos anteriores se producen matrices de melanosoma, la miosina funciona en ambas tambien y las matrices se depositan en la plumas aunque van a ser incoloras pequeñas o deformadas, la produccion de eumelanina es tan baja que es imposible verla a simple vista, solo con microscopios potentes.La ausencia o disminución de pigmento en el cuerpo torna habitualmente a los animales albinos más vulnerables y hace que sean más detectables por sus depredadores por lo que es difícil, pero no imposible, encontrar animales albinos en la naturaleza.
- En los fallow bronce( falow tipo 1); la actividad de la tirosinasa disminnuye parcialmente, con lo cual hay una cantidad menos de produccion de pigmento de calidad
- En los fallow pale( fallow tipo 2); hay una reduccion en la calidad de las matrices que desemboca en la disminución cualitativa de la eumelanina.
-Canela; en esta mutación la eumelanina no se vuelve completamente negra durante la sintesis, produciendose eumelanina de color marron, y las matrices tiene una forma normal.
-Reducciones cuantitativas( cantidad); ; Son los edged, dilute y pastel.
Se deben a un defecto en la distribución( transporte) de la eumelanina y esta relacionado con la presencia o ausencia de los medios de transportes que son;
- miosina
-dendritas de los melanocitos
Los granulos de eumelanoma se desarrollan de manera normal, pero no todos terminan en la pluma. Estas mutaciones no afectan al deposito de eumelanina en la piel.
En dilutes; cuando la eumelanina que se ha formado se deposita en las plumas, se forman varios granulos de eumelanina que se unen uno a otros dando lugar a los macromelanosomas.
-Leucismo; son los arlequines tanto recesivos como dominantes.
Aquí la tirosinasa o el transporte por parte de la miosina no tienen nada que ver, la causa se encuentra en la cresta neural, la migracion de los melanoblastos a la piel o a su entorno, observaremos que en ciertas partes de la piel no hay casi melanocitos, y por esta razon si no hay es imposible que el pigmento se deposite en las plumas de esa zona. Para entender mejor que es la cresta neural en esta pagina al final hay una foto con un embrion de ave en el huevo, ahi podemos visualizar la migracion de los melanoblastos y lo que es la cresta neural, al final de la pagina.
http://www.canariculturaenpuertorico.com/index_files/Page1800.htm
En los arlequines recesivos; hay un defecto en la distribución de las celulas pigmentarias desde la cresta neural( que es el lugar donde se originan lo melanoblastos, celulas percursoras de los melanocitos) como resultado pocos o ningun melanocito llega a la piel, la enzima miosina( recordad que es la que transporta) tiene un funcionamiento normal pero como no hay melanocitos suficientes no se puede pigmentar esa parte de piel y pluma.
En los arlequines dominantes; ciertas partes de la piel se cambian geneticamente de forma que los melanocitos son incapaces de quedarse en la piel o mueren, en esta zona de las no hay matrices depositadas en las plumas y son areas sin pigmentación.
Fuentes;
http://www.cockatielworld.org/genetics.html
Aurora Carranza Roman
FORMACION DE LAS MELANINAS; eumelanina y feomelanina
¿Cómo se forman la eumelanina y la feomelanina?
Este proceso tiene lugar en el interior de los melanosomas (feomelanosomas si contiene feomelanina y eumelanosomas si contienen eumelanina), a partir de la tirosina. Una sucesión de oxidaciones de tirosina catalizada por la tirosinasa conduce a la síntesis de DOPA (dihidroxifenilalanina)para producir, a continuación, un compuesto intermediario común: la dopaquinona.
A partir de este punto, dos vías distintas conducen a la formación de la eumelanina y de la feomelanina;
-Si la enzima cisteina se añade a la dopaquinona obtendremos feomelanina.
- Si la enzima anterior no se une a la dopaquina el proceso de desarrolla completamente, y la enzima miosina vierte los granulos de eumelanina (melanosomas) a la pluma a traves de las dendritas.
Aurora Carranza Roman
PIGMENTOS; PSITACINA Y EUMELANINA EN AGAPORNIS
PSITACINA
Los loros son los únicos pájaros que poseen los pigmentos de la psitácina, estos pigmentos producen las plumas rojas, anaranjadas, y amarillas.La dieta de un loro, por lo tanto, no controla el color de sus plumas.
Algunos autores se refieren a ella como carotenoides pero esta mal expresado ya que los pigmentos de la psitacina ( tambien llamadas psitacofulvinas)son diferentes a los carotenoides.
Los carotenoides son un grupo de pigmentos vegetales liposolubles de color intenso (rojo, anaranjado y amarillo). Los carotenoides se encuentran en todos los alimentos de origen vegetal. Su color, que varía desde amarillo pálido, pasando por anaranjado, hasta rojo oscuro, se encuentra directamente relacionado a su estructura quimica.
Los animales son incapaces de sintetizar carotenoides y deben obtenerlos a través de su dieta (por ejemplo, el color de las canarios rojos, este color se puede intensificar por la adición de ciertos elementos alimenticios a la dieta como pastas especiales).Los pigmentos carotenoides se metabolizan en el hígado, pero el aspecto del pájaro no cambia hasta que las plumas nuevas crecen. Los científicos del comportamiento animal sugieren que el brillo del plumaje coloreado por los carotenoides indica la condición física de un pájaro masculino e influencian la selección del compañero por parte de la hembra
En los loros y por lo tanto tambien en los agapornis, los alimentos no influencian generalmente la intensidad del color. Esto no quiere decir que un pájaro sano con una buena dieta no obtiene colores más vibrantes, pero la buena salud es lo que mejora el brillo, ya que se reflejara mejor la luz en sus plumas, pero no es un cambio de color.
Los agapornis absorben los carontenoides (luteína, zeaxanthin, ß-cryptoxanthin)pero no los fijan a sus plumas, Los carotenoides obtenidos de las frutas, y de las semillas están presentes en la circulación sanguínea de los loros a la hora del crecimiento de la pluma, pero sus plumas se colorean con las psittacofulvinas(psitácina)no con los carotenoides .La psitácina no se encuentra presente en la circulación sanguinea , sino que se sintetiza en el foliculo de donde nace la pluma, asi que el color de los roseicollis red suffusión podria ser debido a una enfermedad o alteracion en el foliculo.
El termino psittacina fue acuñado para hacer claramente la distinción entre los pigmentos encontrados en loros y los lipocromos (carotenoides) encontrados en otros pajaros como por ejemplo los canarios.
La psittacina que va del amarillo al rojo puede estar presente en una forma pura y también en mezcla. Las partes de la pluma que contienen psittacina solamente solo pueden tener un color que va del amarillo claro al naranja y al rojo brillante. Cuando estas plumas también contienen el melanina el color variará a un amarillo marrón o verde oliva oxidado( agapornis nigrigenis) a marrón negruzco o a verde. Si la concentración del melanina es muy alta enmascarará el color de la psitacina
subyacente ( ejemplo cabeza del personata).
MELANINA
Es un compuesto oscuro, un pigmento, que se produce solamente en dos tipos de células;
-en los melanocitos ( que estan en piel y plumas)
-en las células del epitelio pigmentado de la retina (en el fondo de la retina de los ojos de nuestros pajaros).Los colores de los ojos son producto también de la diferente proporción de ambas melaninas(eumelanina y feomelanina) y de la distribución de las células pigmentarias en el iris.
Dado que la melanina debe producirse por la acción de varios enzimas, cuya información genética se encuentra codificada en los genes correspondientes, si alguno de estos genes presenta alguna anomalía( en las mutaciones ocurre esto) el enzima resultante no funcionará o funcionará inadecuadamente, obteniéndose la cesación o disminución de la producción de melanina.
Hay dos tipos de melanina :
-la melanina oscura (negruzca-marronácea), denominada eumelanina
-la melanina más clara (pardo-amarillante-rojiza), llamada feomelanina
La melanina se produce y acumula dentro de los melanosomas.A continuación explicaremos que son;
Melanosomas; son unos orgánulos de la celula, unos sacos membranosos, donde se acumulan y organizan todas las enzimas necesarias para la síntesis de melanina. Dentro de los melanosomas se produce la melanina que, al ser tóxica para la célula debe estar recogida y aislada del resto de orgánulos y substancias celulares. Los melanosomas son unos orgánulos celulares especializados que se forman a partir de los lisosomas del reticulo endoplasmatico de la celula
Fuentes;
http://javarm.blogalia.com/historias/27084
http://www.sciencedaily.com/releases/2005/02/050223123815.htm
http://www.livescience.com/imageoftheday/siod_050223.html
http://researchmag.asu.edu/stories/parrots.html
http://www.public.asu.edu/~kjmcgraw/pubs/TheBiochemist.pdf
http://www.sciencemag.org/cgi/content/abstract/309/5734/617
http://animalscience.ucdavis.edu/research/parrot/
http://classifieds.friendster.com/us/View/47447/15561.html
Aurora Carranza Roman
GENÉTICA BÁSICA
GENÉTICA
Para entender las mutaciones en los agapornis es necesario saber un poco de genética, no es necesario que lo aprendas todo de memoria pues te resultara aburrido e incomprensible, mi intención no es esa, sino que volváis a este apartado para entender conceptos conforme vayáis avanzado en vuestro aprendizaje.
Todos los seres vivos están constituidos por células. Durante la reproducción de las células, las cadenas de ADN que componen los cromosomas se hacen visibles en el centro. A continuación expondré el significado de varios términos usados en genética;
- Fenotipo es la manifestación y expresión del genotipo (de la información genética. Es el color que vemos del pájaro
- Genotipo; es el contenido genético de un individuo, en forma de ADN. Lo que no vemos a simple vista, sino que esta en sus genes.
-Homocigótico; hablamos de un ser homocigótico cuando la combinación es la misma en ambos genes, ( recuerda los cromosomas van de dos en dos, y los genes al estar dentro de los cromosomas también)
-Heterocigótico; cuando la combinación es diferente en ambos genes
-Mitosis; proceso de división celular que produce células genéticamente idénticas, es el fundamento del crecimiento, de la reparación tisular, es decir es la forma mediante la cual los pichones se desarrollan, es la responsable del crecimiento de tegumentos( Plumas, uñas, pico, piel), curación de heridas...
-Meiosis; proceso que comparte mecanismos con la mitosis pero que no debe confundirse con ella, produce células genéticamente distintas y, combinada con la fecundación, es el fundamento de la reproducción sexual.
- Cromosoma; Es el material constituido del ADN que se encuentra en el núcleo de la célula. Los cromosomas contienen genes en un lugar concreto (denominado locus. Se agrupan de dos en dos
- Cromosomas Homólogos: Par de cromosomas integrado por uno proveniente del progenitor femenino y otro proveniente del progenitor masculino. Cromosomas que se aparean durante la meiosis
Como existen cromosomas autosomales y cromosomas sexuales, un gen que se encuentre en el cromosoma Z se le denominara ligado al sexo. Cuando un gen se encuentra en un autosoma decimos que es autosómico.
Para escribir formulas genéticas se usan estos sistemas de determinación
- Sistema de determinación XY: es propio del ser humano y muchos otros animales. ; hembras son XX y los machos son XY
- Sistema de determinación XO: otras especies (peces, insectos, anfibios) que no tienen el cromosoma Y, determinándose el sexo por el número de cromosomas X, macho XO y hembra XX.
- Sistema de determinación ZW: ocurre lo contrario, el sexo masculino es homogamético (ZZ) y el femenino heterogamético (ZW. Es el caso de los agapornis y mariposas.
Nosotros nos vamos a centrar en el sistema de determinación ZW:
En las fórmulas genéticas se escribe ZZ para los machos y ZW para las hembras
MACHO HEMBRA
ZZ ZW
ZZ ZW
.
El cromosoma W es aproximadamente un tercio del cromosoma Z y contiene poca información genética. Hasta hoy, no se han encontrado genes productores del color en este cromosoma W, así que por conveniencia, cuando hagamos cálculos de color lo consideraremos como vacío.
- Gen; un gen es la unidad básica de herencia, es una secuencia de nucleótidos en la molécula de ADN, que contiene la información necesaria para la síntesis de una macromolécula con función celular específica.
El gen es considerado como la unidad de almacenamiento de información y unidad de herencia al transmitir esa información a la descendencia. Los genes se disponen a lo largo de cada uno de los cromosomas.
- Locus ; posición determinada que ocupa cada gen.
- Alelos ;un alelo es una de las formas variantes de un gen en un locus (posición) Diferentes alelos de un gen producen variaciones en las características hereditarias tales como el color sangre.
- Tipos de alelo; los genes pueden aparecer en versiones diferentes, con variaciones pequeñas en su secuencia, denominadas alelos.
Los alelos pueden ser dominantes o recesivos;
-Dominante; Cuando una sola copia del alelo hace que se manifieste el rasgo fenotípico,
-Recesivo; Cuando son precisas dos copias del alelo (una en cada cromosoma del par),
Los alelos múltiples son formas diferentes, expresiones del mismo gen.
- Mutacion; es una alteración o cambio en la información genética (genotipo) de un ser vivo y que, por lo tanto, va a producir un cambio de características, que se presenta súbita y espontáneamente, y que se puede transmitir o heredar a la descendencia.El tipo ancestral o color salvaje de un pájaro es el estándar o el pájaro “completo”.
Tipos de mutaciones ;
- Mutaciones de genes; Son las mutaciones que ocurren al alterar la secuencia de nucleótidos del ADN.
Para entender algunas mutaciones ejemplo ; cinammon- ino. Opalino-ino....vamos a explicar que es un crossing over
Crossing-Over : Proceso que ocurre en la meiosis e incluye la ruptura de un cromosoma materno y uno paterno (homólogos), el intercambio de las correspondientes secciones de ADN y su unión al otro cromosoma. Este proceso puede resultar en un intercambio de alelos entre cromosomas, a través de los cruces pueden producirse nuevas combinaciones de alelos produciéndose un color distinto.La frecuencia con la que esto ocurre se llama frecuencia de cruce.
- Mutaciones cromosómicas
- Mutaciones del genoma
Aurora Carranza Roman
Suscribirse a:
Comentarios (Atom)
